Introduzione alla linguistica generale.
Materiali integrativi al corso di Didattica delle lingue moderne.
di Manuel Barbera (b.manuel@inrete.it).
1. Il linguaggio.
1.5 Gli altri linguaggi (animali ed artificiali) e l'origine del linguaggio.
1.5.0 Impostazione del problema.
Nei paragrafi precedenti abbiamo presentato le caratteristiche
generali per cui un sistema possa essere inteso come "linguaggio" (§ 1.0), ed abbiamo
poi approfondito le principali specifiche di quel particolare linguaggio che č il
nostro. Indagare, ora, sulla natura e l'origine del linguaggio porta inevitabilmente
a domandarsi quali siano, e se vi siano, altri "linguaggi" di fatto esistenti al mondo
oltre a quello umano, alle sue lingue storiche ed alle sue costruzioni artificiali.
Č importante l'impostazione che si dŕ al problema: la domanda, propriamente, č (1)
quel complesso sistema di comunicazione che č il linguaggio umano č specifico solo
dell'uomo o no?, o (2) l'essere forniti di una qualche forma di linguaggio č comune
ad altri (alcuni / molti / tutti ?) organismi viventi? e se sě, ha qualche relazione con
la nozione di "vita" stessa?
I due modi di approccio sono diversi e le risposte che si otterranno sono diverse: la
prima sostanzialmente negativa e la seconda positiva, come vedremo tra poco.
In ogni modo, tutti gli approcci al problema e tutte le "soluzioni" che vi possono dare
partono dai risultati dell'evoluzionismo, una teoria scientifica sull'origine della vita che ha
prodotto risultati straordinari da quando nel 1859 Charles Darwin, con la sua Origin
of Species (ne esiste una utile versione web
commentata e guidata resa liberamente disponibile dalla BBC), la propose ad oggi.
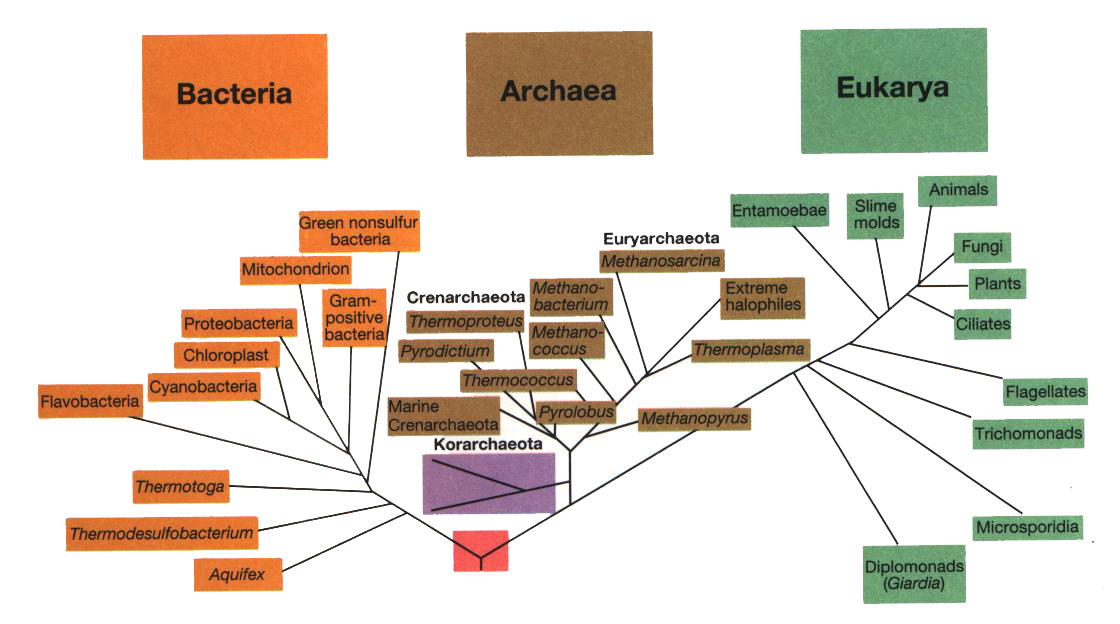
Visto che le scoperte nella filogenesi dei viventi, e quindi i mutamenti
nella tassonomia standard, sono stati e continuano ad essere assai profondi e veloci
(quello che avevo studiato io sui libri del liceo, ad esempio, č giŕ completamente
sorpassato), vi propongo uno schema generale recente, abbastanza consensuale nella comunitŕ
scientifica odierna (anche se in qualche dettaglio, come vedremo, č giŕ sorpassato anch'esso!),
che ci potrŕ essere utile tener presente, non fosse che per renderci conto che il vecchio "regno
animale" (in cui oltre tutto gli "animali superiori", vertebrati sono solo un piccolo piů evoluto
raggruppamento: cfr. la tavola dei Metazoa riprodotta piů indietro)
č solo la punta, esigua ma piů avanzata, dell'iceberg di uno dei tre effettivi regni
del vivente, quello degli eucarioti.
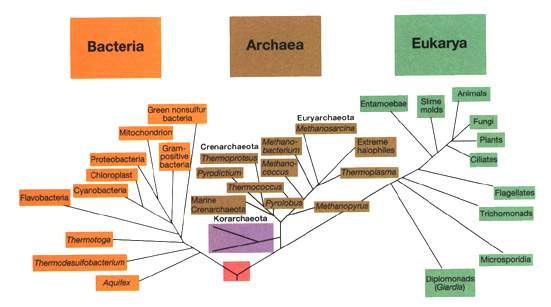
[tav. 1]
La filogenesi del mondo vivente: l'albero universale dela vita.
Riprodotto dal classico manuale Brock. Biology of Microorganisms, 9th edition
by M. T. Madigan, J. M. Martinko and J. Parker, Upper Saddle River (New Jersey),
Prentice-Hall, 2000. L'albero č derivato dalla sequenziazione del RNA 16S o 18S,
basandosi sui dati del Ribosomal Database Project.
La distanza evolutiva tra due gruppi di organismi č proporzionale alla distanza
cumulativa tra la fine del ramo ed il nodo che congiunge i due gruppi. Č disponibile
anche una versione grande della tavola.
1.5.1 I linguaggi artificiali.
La domanda che abbiamo formulata nel paragrafo precedente verte
naturalmente sui linguaggi che potremmo dire "naturali". Esiste infatti un grande
numero di linguaggi "artificiali", costruiti cioč dall'uomo. Tutti sono, dunque, di natura
secondaria e dipendono in modo indiretto dal linguaggio naturale umano. Il loro studio
non ci illumina sulla natura e l'origine del linguaggio, ma č molto interessante
tanto per ragioni pratiche (a chi non piacerebbe sapere programmare al computer?),
sociali (nessuno metterebbe in dubbio l'utilitŕ del linguaggio gestuale per i muti),
scientifiche (non potremmo parlare di fisica senza il linguaggio della matematica) e
metodologiche (un linguaggio artificiale č come un modello in vitro su cui
condurre esperimenti in condizioni assolutamente controllate). Nonostante l'evidente
interesse dell'argomento, dati i nostri scopi, non potremmo approfondirlo molto, e
ci accontenteremo di una breve rassegna dei "tipi" principali, facendo riferimento,
come bussola, alla tavola riassuntiva
posta in fondo a questo capitolo.
Che vi siano molti linguaggi informatici o di programmazione,
da bravi utilizzatori di computer, lo sappiamo tutti, anche se poi magari non ne
conosciamo davvero nessuno ("non sappiamo programmare). Lo
sviluppo
di linguaggi di questo tipo accompagna tutta la storia dell'informatica, anche se si suole di
solito fissare al 1946 la data di nascita del primo di essi, il Plankalkul (č stato sviluppato
da un tedesco, Konrad Zuse). Ne sono ormai stati sviluppati moltissimi (se sei interessato
ne puoi vedere un elenco abbastanza ampio con i principali link disponibili per ognuno
su dmoz,
od una bibliografia
piů selettiva e ragionata) e le loro caratteristiche sono
estremamente variabili a seconda degli scopi per i quali sono stati introdotti.
Molte dei tratti identificativi del linguaggio umano non sono, cosě,
particolarmente diagnostiche: ad es. i "segni" di cui sono costituiti hanno una
diversa materia dell'espressione (unitŕ di impulsi elettronici vs. immagine acustica)
ed una diversa forma dell'espressione (codici binari vs. opposizioni distintive piů
articolate) e di fatto usano un diverso canale, articolazione e composizionalitŕ sono di solito presenti (ma in misura
variabile), vale sempre la ricorsivitŕ ma la dipendenza dalla struttura č piů spesso
evitata, si tratta sě di "lingue storiche" distinte e stabilite convenzionalmente,
ma che non cambiano naturalmente nel tempo (sono semmai sostituite da "versioni" nuove)
e non sono "socialmente" radicate, fanno tutte uso di "strutture specializzate" (memoria
elettronica, RAM o disco) che perň non sono "naturali". Le categorie pertinenti
per una classificazione dei linguaggi di programmazione sono semmai altre (ad es.
interpretati?, compilati?, ad oggetti?, ecc.) nel cui dettaglio non possiamo entrare (vedi
ad es. le "categorie" identificate nella pagina di dmoz linkata sopra).
Analogo discorso va fatto per i linguaggi logici, che vanno
dalla classica logica proposizionale di primo ordine ai piů complicati sistemi di logica
modale (ad esempio "S5") od a piů valori di veritŕ (come quella di Lukasiewicz) alle
piů recenti "fuzzy logics". Naturalmente qui stiamo parlando dei sistemi di logica
storicamente sviluppati, non del problema delle condizioni filosofiche di ogni
logica (i fondamenti generali della logica), problema decisamente troppo grosso (e parzialmente
fuori tema) per un corso di filosofia. A chi fosse interessato a "capire" i problemi
della logica potrei consigliare l'ottimo Susan Haack, Philosophy of Logics, Cambridge
University Press, 1978; per invece "imparare" fattivamente la (una) logica quasi tutti
i molti manuali disponibili possono andar bene.
Vi sono poi anche molte lingue inventate, create ossia deliberatamente
a tavolino da qualcuno per scopi ludici (ad esempio il klingon
creato per l'affabulazione cinematografica di Star Trek)o pratici (ad esempio
l'esperanto). L'invenzione puň partire da zero (o quasi, come nel caso dell'
esperanto
creato nel 1887 dal medico polacco L. L. Zamenhof, 1859-1917) o da stati di lingua preesistenti.
Se il linguaggio inventato diventa la madre lingua di qualche parlante cessa di essere "artificiale"
e prende a svilupparsi come una qualsiasi lingua naturale (naturalmente vi possono essere molte
fasi transizionali tra i due stati). Di solito questo č avvenuto solo con linguaggi (a) non creati
dal nulla e (b) che si rivolgevano a comunitŕ ben definite: č il caso ad es. dell'ebraico
moderno, diventato reale lingua della popolazione israeliana, ma creato "a tavolino"
riportando in vita il defunto ebraico biblico (in diversa misura č il caso anche del
tedesco moderno e dell'italiano stesso: provate a riflettere perché).
Una categoria piů sfuggente, e per la quale non esiste una etichetta
standard, č quella che chiamarei dei metalinguaggi funzionali semplificati ("MFS").
Sono propriamente sistemi per ricodificare in modo tendenzialmente univoco anche se
piů o meno semplificato una lingua (s1) che fa uso di un derminato canale materiale
(c1: tipicamente i suoni del linguaggio umano) in una altra "lingua" (s2: un metalinguaggio,
dunque) che fa uso di un diverso canale materiale (c2). La ricodifica, di norma,
non investe la struttura della lingua stessa (la langue) ma solo la sua espressione
materiale: non abbiamo dunque la creazione di una vera nuova lingua, ma solo una diversa
espressione della lingua stessa: nella tavola riassuntiva,
dunque, i MFS presenteranno caratteristiche "ereditate" dal linguaggio umano che
sottintendono, fatta salva la diversitŕ nel canale utilizzato, e le semplificazioni
effettuate. Alcuni esempi tipici possono essere il cosidetto "American Sign Language" o
ASL
usato per i muti, il MORSE che ricodifica in una sequenza di impulsi i fonemi (in realtŕ
i grafemi ...) di una lingua qualsiasi, o, all'estremo, i sistemi di scrittura medesimi
(sui quali torneremo piů eplicitamente in seguito).
L'ultima classe di linguaggi fabbricati dall'uomo che prenderemo
in considerazione sono quelli che potremmo chiamare codici di segnali:
la loro principale caratteristica risede nel particolare tipo di segni di cui sono
costituiti, che sono dei "segnali" nel senso di ascendenza husserliana che avevamo
introdotto nel § 1.3.5), ossia dei segni (in genere poco complessi) in cui l'aspetto materiale
č assolutamente prevalente. Si tratta, per intenderci, di linguaggi come quello del
semaforo o quello dei dei segnali stradali. Anche per altri aspetti questi sistemi
si diversificano dal linguaggio umano, come si puň vedere dalla solita
tavola riassuntiva: ad esempio,
per la ridotta composizionalitŕ, la limitata ricorsivitŕ, ecc.
1.5.2 La ricerca del simile all'umano: 1. Primati.
Riprendendo il discorso impostato nel § 1.5.0, e prendendo
il problema secondo la prima formulazione, la prima idea che puň venire per
affrontarlo č quella di andare a vedere come stiano le cose negli animali piů
simili evoluzionisticamente all'uomo.
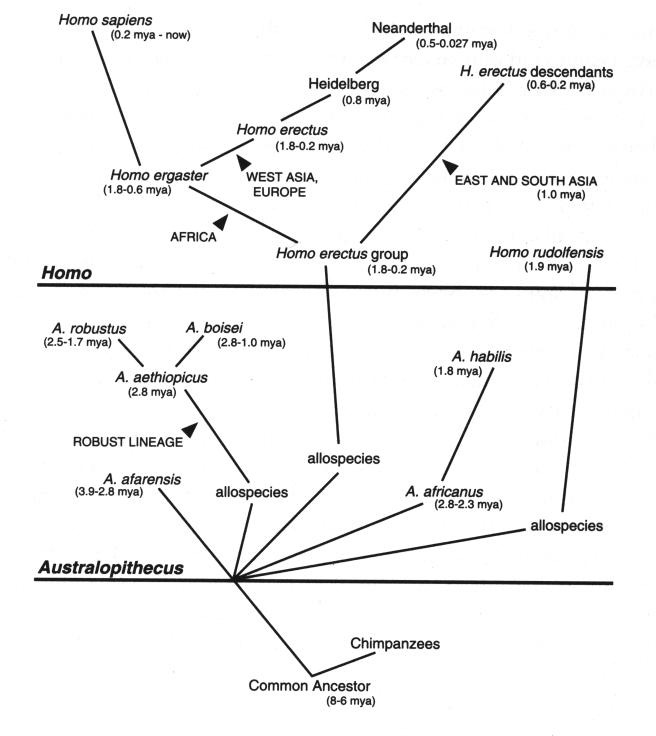
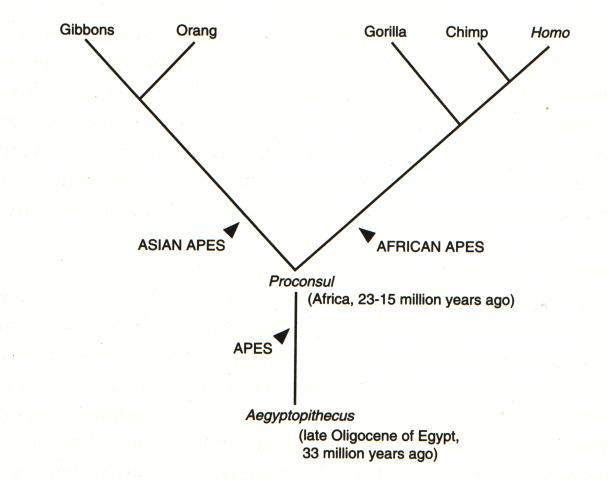
La precisa filogenesi della specie homo sapiens non č chiara in tutti i
dettagli, dato che molto dipende dalle frammentarie attestazioni paleontologiche,
anche se nelle sue grandi linee puň riassumersi nel modo seguente:
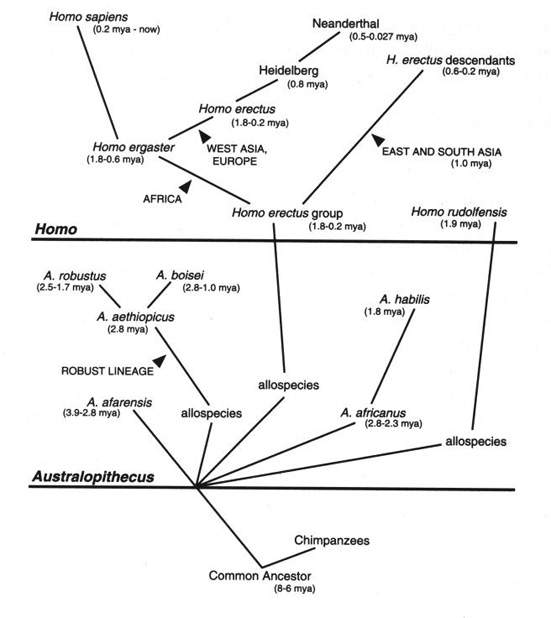
[tav. 2]
La filogenesi dell'uomo: l'albero degli umani.
Riprodotto da Ernst Mayr, What Evolution Is, New York,
Basic Books, 2001. L'albero rappresenta solo l'ipotesi piů probabile sulla filogenesi
dell'uomo, e non tiene conto dei fossili descritti dopo il 1990.
La connessione dell'uomo con le scimmie, comunque, argomentata
giŕ da Darwin nel suo "scandaloso" The descent of man, and selection in relation to sex (1870-1, poi
1874 ed ultima revisione 1877), č ormai del tutto chiarita nelle sue linee filogenetiche,
ossia schematicamente (cfr. invece la tassonomia completa
dei primati della Primate Infonet del Wisconsin; questa č invece una utile
tabella riassuntiva):
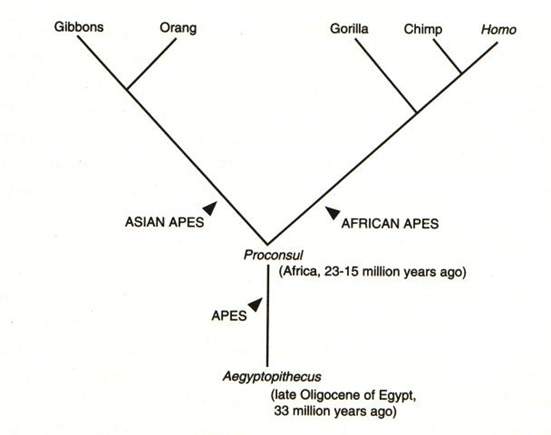
[tav. 3]
La filogenesi degli ominidi (phylum: Vertebrata, [...], ordo: Primates, subordo: Anthropoidea,
infraordo: Catarrhini, superfamilia: Hominoidea [apes], familia: Hominidae):
l'albero della famiglia "Hominidae".
Riprodotto da Ernst Mayr, What Evolution Is, New York, Basic Books, 2001.
Certo, prima di Darwin, propriamente, il problema non si poteva porre
in quanto tradizionalmente (opinione, se vogliamo, influenzata dalla religione, ma
asseverata anche da pensatori come Cartesio!) gli animali "non hanno un anima" (e
quindi sentimenti, linguaggio, ecc.), anche se "ingenuamente" nessun felice proprietario
di animali da compagnia ha mai dubitato dei sentimenti del proprio cane o dell'intelligenza
del proprio gatto. Ma dopo Darwin lo studio perlomeno delle scimmie, o meglio
sugli scimpanzé (i nostri cuginetti piů prossimi) era inevitabile.
I primi, sperimentali, studi compiuti sulle scimmie, tuttavia, fornirono solo
la sconsolante constatazione che non erano capaci di usare un linguaggio proposizionale,
ossia di parlare (cfr. Wolfgang Koehler, Zur Psychologie der Schimpansen,
in "Psychologische Forschung" I (1921)), e ribadendo che erano limitate ad un "linguaggio
emozionale", gestuale e non strutturato. Un punto che non veniva colto, naturalmente,
era che semplicemente non erano predisposte alla fonazione, non possedendo adeguati
apparati articolatori. I primi esperimenti "moderni" sono dovuti a Robert Mearns Yerkes
(1876-1956), uno psicologo della Pennsylvania famoso soprattutto per avere introdotto
per la prima volta test psicologici nella reclutazione dei militari dell'esercito
americano in occasione della prima mondiale, che per primo concluse che «there is
abundant evidence that various other types of sign process than the simbolic [intendi:
"segno linguistico"] are of frequent occurrence and function effectively in the Chimpanzee»
(da Yerkes & Nissen, Pre-linguistic Sign Behaviour in Chimpanzee, in "Science"
LXXXIX, p. 587 cit. in Ernst Cassirer, An Essay on Man. Introduction to a philosophy
of human culture, New Haven - London, Yale University Press, 1992 [1944], p. 30;
cfr. anche il consuntivo delle sue ricerche: R. M. Yerkes, Chimpanzees. A Laboratory
Colony, New Haven, Yale University Press, 1943), sgombrando cosě il campo dalle vecchie
questioni sul "non-linguaggio" emozionale.
Un tipo di ricerca piů mirata (invece di vertere solo sul comportamento,
come usuale in anni di behaviourismo dominante) puň essere quello di individuare
la presenza (fisiologica) di strutture specializzate atte a "sostenere" un linguaggio.
Esperimenti di questo tipo diedero risultati incoraggianti anche su scimmie piů
periferiche rispetto alle Hominoidea, come gli uistiti, le "squirrel
monkeys" dell'America del Sud. In particolare, sono stati individuati alcuni neuroni
nella corteccia uditiva che rispondono selettivamente solo ad alcuni suoni specie-specifici:
si tratterebbe, ossia, di neuroni che in qualche modo individuano
i tratti semantici nei suoni, selezionando di attivarsi solo in presenza di suoni
portatori di significato.
Un altro approccio proficuo č stato quello di indagare il fronte
dell'apprendimento, come giŕ aveva fatto Yerkes, dato che l'acquisizione spontanea naturale
e l'apprendimento in contesto "sociale" č pur sempre una caratteristica saliente del linguaggio.
Le esperienze piů interessanti in questo campo sono state fatte ancora una volta
non direttamente sugli Hominoidea ma bensě sui cercopitechi ("vervet monkeys",
appartenenti ad una superfamilia cognata agli Hominoidea) dell'Ambroseli
National Park in Kenya, studiati da Peter Marler e colleghi della Rockefeller University,
e di cui Dorothy L. Cheney - Robert M. Seyfarth, How Monkeys See the World,
Chicago and London, University of Chicago Press, 1990 fornisce una ricca trattazione ed un quadro stimolante.
I cercopitechi, tra le loro varie vocalizzazioni (cfr. soprattutto i capitoli 4 e 5
di Cheney - Seyfarth 1990 cit.) dispongono di tutta una serie di segnalazioni per l'avvistamento di
predatori, principalmente (a) per i leopardi, (b) per le aquile, (c) per i pitoni, piů
alcuni altri come (d) per i babbuini e (e) per gli umani sconosciuti. Le segnalazioni
studiate sono le prime tre (a-c), che funzionano tutte anche con registrazioni riprodotte. La cosa
piů interessante č che i piccoli riconoscono sě i tre segnali, ma ne fanno un uso
meno specifico, sovraesteso (ad es. usano la segnalazione per l'aquila anche per altri
uccelli, a volte addirittura non pericolosi): solo dopo una fase di acquisizione
relativamente lunga "imparano" il corretto impiego; altrettanto interessante č come
alcuni gruppi di cercopitechi abbiano sviluppato un segnale di avvertimento per
un pericolo (l'uomo) "nuovo" nel loro originario ecosistema.
La maggior parte degli esperimenti piů rilevanti condotti con gli
scimpanzé dopo quelli di Yerkes sono tutti partiti dal presupposto di non usare
direttamente una lingua "fonica" naturale umana, ma di passare attraverso un qualche
tipo di MSF (ricordate la categoria che abbiamo illustrato poco sopra?) come l'
"American Sign Language" (ASL).
Le prime esperienze da riferire in questo settore sono quelle condotte da Beatrice
ed Allen Gardner che, presso l'Universitŕ del Nevada, hanno "educato" la scimpazé
Washoe. I Gardner hanno cercato di allevare Washoe come una bambina, vivendole
insieme ed usando l'ASL anche tra di loro, ma adottando una tecnica di allenamento a
base di remunerazioni per gli usi corretti del linguaggio (a differenza di quello che
accadrebbe per un bambino). Washoe, in queste condizioni, imparň un repertorio di circa
100 segni che riusciva a combinare in "sintagmi" di massimo cinque costituenti, e, pur
non avendo appreso la lingua in modo spontaneo (ma grazie ad un sistema di remunerazione)
ne č giunta a fare un uso relativamente spontaneo. Siamo ancora abbastanza lontani dal linguaggio
umano, ma c'č giŕ qualcosa ...
Un'altra scimpanzé, Sarah, allenata da David Premack dell'University of California
at Santa Barbara con un sistema di oggetti codificanti parole, era invece giunta ad
imparare un repertorio di parecchie centinaia di segni, che tuttavia usava solo in
situazioni di test, laddove Washoe usava il suo piů limitato repertorio anche
in condizioni relativamente libere.
Un'ultima esperienza, che si rivelerŕ in seguito foriera di nuovi fecondi spunti per
la ricerca, č stata quella dell'educazione di un gruppo di scimpanzé da parte di Sue
Savage-Rumbaugh e Duane Rumbaugh allo Yerkes Primate Center di Atalanta (Georgia). I
risultati ottenuti furono in tutto simili a quelli conseguiti con Washo e Sarah, e
comunque gli scimpanzé non reagivano al linguaggio parlato umano ma solo a quello
simbolico. La novitŕ era perň proprio in quest'ultimo: si trattava da un linguaggio,
sempre di tipo MSF, costituito da simboli geometrici rappresentati su una larga tastiera,
lo Yerkish (cosě chiamato in onore di Yerkes, cui era anche intitolato il centro
presso il quale vennero svolte le ricerche). Lo Yerkish si č rivelato un "buon" MSF,
particolarmente utile anche nel caso di persone con gravi forme di autismo o con serie
lesioni cerebrali: le tastiere Yerkish costruite dai Rumbaugh hanno contribuito
a migliorare la vita a moltissime persone svantaggiate. Il che č molto bello, si dirŕ,
ma non sposta di nulla il problema di cui ci stiamo occupando. Ed infatti la storia non
finisce qui.

[tav. 4]
Uno scimpanzé mentre usa la tastiera yerkish. Immagine tratta dal sito italiano sui bonobo.
Lusingati dai successi intrinseci dello Yerkish, ma preoccupati dello
scarso progresso ottenuto con gli scimpanzé, i Rumbaugh ebbero l'idea di provare con i
bonobo (Pan paniscus, impropriamente noti anche come scimpanzé pigmei. Sui
bonobo c'č un bel sito italiano, con
affascinanti fotografie), che piů recenti ricerche filogenetiche avevano evidenziato come
ancora piů vicini all'uomo dello stesso scimpanzé proprio. Allevarono pertanto il bonobo
Kanzi, educandolo insieme alla madre Mattata. Da 6 mesi (etŕ corrispondente a quella
della lallazione umana) a 2 anni e mezzo Kanzi fu esposto
all'apprendimento dello Yerkish insieme alla madre, dalla quale venne poi separato
ed allevato da umani (i Rumbaugh medesimi). Un fatto importante č che lo Yerkish fu
insegnato a Kanzi in modo il piů possibile naturale, senza ricompense e sessioni
distinte di training. I risultati furono subito eccezionali: Kanzi usava la tastiera
spontaneamente (a differenza degli scimpanzé), e, quando aveva un vocabolario ancora
limitato a sole 35 parole, avvenne che mentre i ricercatori parlavano inglese tra di loro
riconobbe spontaneamente una parola e la riprodusse sulla sua tastiera: Kanzi, in
altre parole stava imparando l'inglese. Č stato questo il primo caso che si sia registrato
di una indiscutibile interazione linguistica tra uomo ed animale: l'esperimento,
tra l'altro č recente ed in corso, e tuttora Kanzi non cessa di stupire. I dati piů
recenti sono quelli forniti in Duane M. Rumbaugh - David A. Washburn, Intelligence
of Apes and Other Rational Beings, New Haven and London, Yale University Press,
2003 (cfr. soprattutto il § 4, pp. 128-143) e sono affatto sorprendenti: Kanzi ormai
comprende centinaia di lessemi e frasi di moderata complessitŕ (del tipo "Go to
location X and get the object Y", "Get the carrot that's in the microwave", ecc.)
nella lingua inglese, che poi sa anche (in misura minore: non diversamente da come
avviene l'apprendimento negli umani) usare naturalmente (ricodificati nel metalinguaggio
non verbale).

[tav. 5]
Kanzi con Sue Savage-Rumbaugh. Immagine tratta dal sito italiano sui bonobo.
1.5.3 La ricerca del simile all'umano: 2. Uccelli, Insetti.
Le esperienze che abbiamo esaminato fino ad ora evidenziano che
alcune specie animali (almeno i bonobo e gli scimpanzé, evoluzionisticamente assai vicine
alla nostra) pur non possedendo in natura un linguaggio strutturato come l'umano
(o possedendolo con le "caratteristiche umane" - articolazione, ricorsivitŕ ecc. - assai ridotte),
sono tuttavia in grado di impararlo naturalmente e di utilizzarlo rudimentalmente:
Kanzi, almeno, ne č sicuramente in grado. Non possiamo, quindi, essere ormai recisi
come Yerkes cinquanta anni fa nel pensare che prima dell'uomo ci si trovi sempre e
solo in una fase assolutamente prelinguistica.
Sorge a questo punto una domanda spontanea: il linguaggio umano (che chomskyanamente
č niente meno che un organo biologico, sottoposto a tutte le leggi dell'evoluzione),
dato che l'abbiamo finora visto compatibile solo con le scimmie antropomorfe filogeneticamente
piů prossime al Homo sapiens, sarŕ una struttura genetica emersa gradualmente nella
filogenesi dei Primati, od elaborata in modo parzialmente indipendente in vari
rami a partire da qualche caratteristica piů generale? Per rispondere
alla domanda bisogna andare a guardare cosa succede anche al di fuori dei Primati
Tra i mammiferi, comportamenti chiaramente "linguistici" sono stati
riportati soprattutto per i delfini ed i pipistrelli, ma con le "caratteristiche umane"
ancora piů attenuate e con scarse prove di effettiva intercomunicazione (che non
siano, ossia, casi di comunicazione non linguistica come quelli del cane istruito
ad andare a prendere le ciabatte) tra uomo e animale (anche se per i delfini
esistono alcune esperienze significative anche in questo senso).
Sorprendentemente esempi molto piů convincenti sono stati riportati
tra gli uccelli, in particolare tra i pappagalli (Psittacus erithacus),
dei quali č assolutamente da sfatare la proverbia le stupiditŕ e capacitŕ meramente
imitativa.
Notevole č soprattutto il caso del pappagallo Alex, allenato a partire dal 1977
da Irene Pepperberg, i cui risultati (l'esperienza credo sia tuttora in corso) sono
stati da lei pubblicati in una serie di contributi a partire dal 1981 (Functional
Vocalization by an African Grey Parrott (Psittacus Erithacus), in "Zeitschrift
für Tierpsychologie" LV (1981) 139-160). Un utile riassunto č fornito da Felice
Cimatti, Mente e linguaggio negli animali. Introduzione alla zoosemiotica cognitiva,
Roma, Carocci, 1998, § 3.1.6, pp. 130-133. Alex nel 1990 era in grado di comprendere
e produrre «circa ottanta parole inglesi, con cui č in grado di riferirsi a oggetti,
forme e materiali» (Cimatti, cit. p. 130): «Alex usa [cioč] le parole inglesi che
č in grado di comprendere per identificare gli oggetti, per chiederli e per rifiutarli»
(Cimatti, cit. p. 131). Alex «č capace di rispondere a 'semplici'
domande come "what colour is [designated object]" [...], "what object is [designated
shape]"» (Cimatti, cit. p. 130); «č anche in grado [...] di usare "no" per non ubbidire
ad una richiesta degli sperimentatori» (Cimatti, cit. p. 132) ed č in grado di contare
fino a dieci oggetti.
K: OK, Alex; cominciamo. Cos'č questo? (in mano ha un bottone di legno)
ALEX: Bottone [di] legno
I: Ha detto; "bottone di legno"
K: Giusto, qui c'č un bottone di legno, Alex. Sei in gamba (offre ad Alex l'oggetto)
ALEX: (prende l'oggetto con il becco, ma subito dopo lo lascia cadere) Voglio tappo
K: OK, ecco il tappo (lo dŕ al pappagallo)
ALEX: (gioca con il tappo per qualche tempo)
K: Ora basta, Alex. Dammi il tappo (allunga la mano verso Alex, che gli dŕ il tappo)
Che cos'č questo? Che forma ha? (ha in mano un pezzo di legno rosso triangolare)
ALEX: Ango..lo legno [Alex indica gli oggetti triangolari con "three corner"]
I: Credo che abbia detto "angolo legno"
K: Alex, che forma ha? Parla chiaramente!
ALEX: Tri-angolo legno
I: "tri-angolo legno"
K: Giusto, la forma č tri-angolo legno (Dŕ ad Alex il pezzetto di legno)
ALEX: (lo afferra, ne morde un angolo, lo lascia cadere)
Voglio nocciola
(K gli dŕ una noce, che Alex mangia)
K: Guarda, cos'č questo? (mostra ad Alex un pezzetto di cuoio grigio)
ALEX: Voglio nocciola
K: No noce ... prima dimmi che cos'č questo?
ALEX: Grigio cuo....
I: "grigio cuoio"
K: Bravo pappagallo ... qui c'č il cuoio grigio (lo offre ad Alex)
ALEX: (rifiuta il cuoio) Voglio nocciola
[tav. 6].
"Dialogo" tra il pappagallo Alex e due sperimentatori, "I" (Irene Pepperberg) e
"K" pubblicato da Irene Pepperberg nel 1988 (An Interactive Modelling Technique
for Aquisition of Communication Skills: Separation of "Labelling" and "Requesting"
in a Psittacine Subject in "Applied Psycholinguistics" IX (1988) 59-76. «I č voltata rispetto al pappagallo
e a K, e ascolta quello che dice l'uccello senza vedere quel che K gli ha chiesto
di nominare, in modo da non farsi influenzare - nel riconoscere le parole di Alex -
da quanto si aspetterebbe che dicesse se avesse potuto vedere l'oggetto in questione»
(Cimatti, cit. p. 131). Riprodotto da Felice Cimatti, Mente e linguaggio
negli animali. Introduzione alla zoosemiotica cognitiva, Roma, Carocci, 1998,
p. 131-2, che lo riprende e traduce in italiano (l'esperimento originale č in inglese)
da Pepperberg 1988 cit., pp.69-70.
Quello che č «ancora piů interessante č il fatto che molte delle richieste di Alex
si riferiscono ad oggetti non presenti nel momento in cui li richiede, né che, peraltro,
sono stati nominati precedentemente dagli sperimentatori (Pepperberg 1988, p. 70).
Questo significa che Alex usa spontaneamente il rudimentale linguaggio
che gli č stato insegnato» (Cimatti, cit. p. 133).
Certo, nonostante la ben diversa collocazione nella filogenesi dei vertebrati, gli esperimenti
condotti su Alex sembrano aver dato risultati dello stesso ordine e grado di quelli
ottenuti con alcuni primati (scimpanzé), cosa che farebbe propendere per l'ipotesi
che si tratti di evoluzioni parzialmente indipendenti (e non, pertanto di una "sinapomorfia"
individuante i mammiferi, come direbbero i biologi) a partire da una comune "predisposizione"
linguistica.
Ma fuori dai vertebrati cosa possiamo trovare?
L'esempio piů noto, fuori dai vertebrati, si ritrova negli insetti
(una classe popolosissima di Arthropoda, cfr. la tavola
tav. 1 nel quarto capitolo),
ed č quello delle api (Apis mellifera), noto giŕ almeno dall'Ottocento. Anzi, si puň dire che molto
spesso, specie nella letteratura generativa, la "danza delle api" č l'unico tipo
di linguaggio animale menzionato. Noi ne diremo molto poco, dato che anche sul Graffi
- Scalise, che in questo non fa eccezione, se ne parla ampiamente.
L'unica cosa che ci limiteremo ad osservare č come si tratti in questo caso di
un "linguaggio" radicalmente diverso dall'umano: le api bottinatrici (le addette
all'aprovvigionamento dell'alveare) comunicano tra loro posizione, distanza e, piů
grossolanamente, quantitŕ dei fiori fonte di polline e nettare, eseguendo particolari
evoluzioni in volo ("danze delle api"). Delle due varietŕ di danze č soprattutto
la "danza ad otto" ad essere interessante, e ne č illustrato il funzionamento nella
tavola seguente.
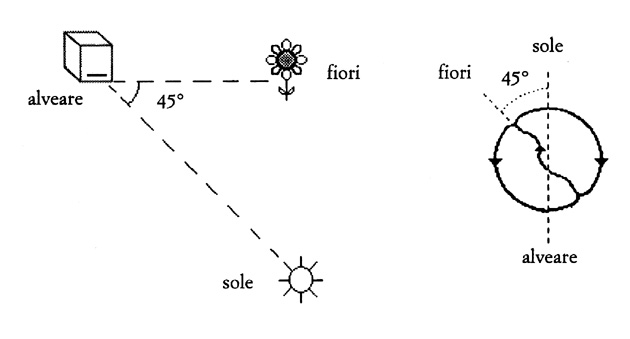
[tav. 7]
La danza ad otto delle api bottinatrici; riprodotta da Felice
Cimatti, Mente e linguaggio negli animali. Introduzione alla zoosemiotica
cognitiva, Roma, Carocci, 1998, p. 66. La posizione dei fiori rispetto all'alveare
viene indicata dall'asse del percorso ad otto della danza, che riproduce l'angolo
dei fiori rispetto alla linea alveare-sole (in questo esempio i fiori sono a 45°
a sinistra del sole rispetto all'alveare); «la distanza dall'alveare del luogo dove si trova il cibo
viene rappresntata mediante il numero di evoluzioni rotatorie complete - cioč di
percorsi a otto - in una data unitŕ di tempo» (Cimatti, cit. p. 65); «La ricchezza
della fonte di cibo, infine, č segnalata dal vigore con cui le bottinatrici si impegnano
nella danza» (Cimatti, cit. p. 67).
Sulle api e sulla loro danza il testo storico di riferimento č ancora
Karl [Ritter] von Frisch, The Dance Language of Bees, Cambridge (MS),
Harvard University Press, 1967, un classico dell'etologia; piů recente ed a vasto raggio č anche
James L. Gould - Carol Grant Gould, The Honey Bee, New York, W. H. Freeman, 1988.
Per quanto si tratti sicuramente di una comunicazione di tipo linguistico,
č evidente la assolutŕ diversitŕ dal linguaggio di tipo umano (e non ne sorprende
la frequente menzione dai generativisti, che di solito vogliono restringere il "linguaggio"
al solo "linguaggio umano").
Prendendo il problema da questa angolatura (ricerca del simile all'umano) si č
in effetti raggiunto probabilmente il fondo della botte. Sarŕ forse allora tempo
di affrontarlo dall'altra angolazione, rinunciando alla impostazione antropocentica
della domanda, che diventa "l'essere forniti di una qualche forma di linguaggio č comune
ad altri (alcuni / molti / tutti ?) organismi viventi? e se sě, ha qualche relazione con
la nozione di "vita" stessa?". Nei capitoli seguenti cercheremo di rispondervi.
1.5.4 Strutture fisiologiche e memoria: Cnidaria e Mollusca.
L'approcio sperimentale che abbiamo introdotto per gli uistiti,
(mirare all'individuazione di strutture specializzate atte a "sostenere" un linguaggio,
correlandole poi a dati comportamentistici) opportunamente adattato si č rivelato utile
anche per la ricerca in organismi inferiori. Si potrebbe, ossia, veder se un organismo
dispone di strutture per rilevare stimoli ("recettori") e quindi imparare ("memoria": e
si ricordi che lo schema
fornito precedentemente vale perlomeno per tutti i mammiferi) dall'esterno. Sappiamo
che i semplici stimoli non sono ancora propriamente linguistici (nel senso perlomeno
che non sono intenzionali), ma sono pur sempre una struttura simbolica, sono piů facili
da rilevare sperimentalmente, e sono comunque in qualche modo un presupposto inevitabile
ad ogni forma di linguaggio: il riscontrarli, quindi, non risolverŕ completamente il
nostro problema, ma ci dice senz'altro qualcosa di molto significativo.
Esperimenti in questo senso sono stati condotti soprattutto
su Cnidaria (anemoni di mare), meglio noti con la vecchia e meno accurata etichetta di "celenterati",
e Mollusca (lepri di mare). Gli cnidari, in particolare, sono organismi che si collocano nel
ramo piů basso e primitivo degli animali veri e propri (Eumetazoa, potete
ricontrollare la tavola che vi avevo fornito),
al di sotto del quale, con l'esiguo ponte dei Porifera (le spugne) e pochi altri
organismi, si passa dal pluricellure all' unicellurare. I molluschi (riportatevi sempre
alla stessa tavola), uno dei phyla piů
numerosi di tutti i metazoi, sono appena di poco piů evoluti (hanno giŕ la simmetria
bilaterale, ad es.). Entrambi i phyla, comunque, come tutti i pluricelluri,
dispongono di strutture specializzate, tra le quali, come tutti gli animali, anche un
sistema nervoso, per quanto primitivo e costituito solo da una rete di gangli nervosi
(un sistema nervoso centralizzato, ricordo, compare solo nel gruppo dei Chordata).
Le Aplysia (aplisie
o lepri di mare, un tipo di lumachine marine), invertebrati (molluschi) dell'ordine
Gastropoda, sono state studiate (cfr. la pagina dell'
APLYSIA invertebrate neurophysiology database project)
in America dapprima da Eric Kandel e colleghi della Columbia University, con l'intento di individuare
neuroni specializzati (analogamente all'esperimento riportato per le scimmie scoiattolo).
Questo tipo di studi č stato proficuamente continuato da Donald
Gardner,
che ha costruito delle reti neurali (non "simulate" come quelle di McClelland di cui
abbiamo riferito nel § 1.4.6) usando appunto i neuroni specializzati che erano
stati identificati (piů precisamente usando «a population of synaptically interconnected
identified neurons in ganglia of the sea snail Aplysia californica», cito dal suo sito).
Gli studi sulle Actinia (attinie od anemoni di mare; cnidari), condotti in Russia dal
neuroscienziato Eugene Sokolov, hanno mirato invece a identificare anche negli cnidari
una memoria strutturalmente simile a quella dei mammiferi, concentrandosi su quel
particolare tipo di memoria procedurale che č la habituation o 'assuefazione'
(cfr. lo schema dei tipi di memoria
a lungo termine): l'attinia quando č toccata contrae i suoi tentacoli, ma, se l'esperienza
viene ripetuta piů volte a distanza di tempo ravvicinato e senza danno, contrae sempre meno.
Il fenomeno, inoltre, Sokolov ha ben chiarito che non dipende semplicemente da una reazione
meccanica e fisiologica come la fatica muscolare (a qualcuno forse l'obiezione sarŕ
venuta in mente ...), in base a confronti con quanto avviene in altri animali.
Per tirare le conclusioni, questo tipo di ricerche hanno dimostrato
che la "memoria" č giŕ presente nei piů primitivi organismi (animali) dotati di
un sistema nervoso. Un requisito essenziale per la esistenza di un linguaggio, pertanto,
č stato dimostrato al di lŕ di ogni possibile dubbio. Ma non che esso vi sia, si
dirŕ.
1.5.5 Verso l'unicellulare: Acrasiales ed eucarioti.
Se facciamo un ulteriore passo indietro nell'albero filogenetico,
affrontando finalmente un organismo unicellulare, scopriremo perň (forse con stupore ...)
che una effettiva comunicazione, con caratteristiche che potremmo ben intendere come
"linguistiche", č stata di fatto descritta. Queesto importante esempio di comunicazione
in organismi eucariotici molto primitivi viene dagli studi di Bonner (1947 e 1959)
e Shaffer (1953, 1956 e 1957) su un particolarissimo gruppo, le Acrasiales; si tratta
di esperimenti "vecchi" e ben noti, ma che forse, essendo stati condotti in anni
in cui la ricerca animale era ferma all'impostazione di Yerkes, sono sfuggiti all'attenzione
dei filosofi del linguaggio e dei linguisti generali. La loro prima valorizzazione dal
punto di vista che ci interessa č probabilmente avvenuta in campo biosemiotico, con
l'importante volume Zoosemiotics (1968) di Thomas A. Sebeok (trad it. Zoosemiotica.
Studi sulla comunicazione animale, Milano, Bompiani, 1973), studioso la cui importanza
abbiamo giŕ sottolineato.
Per potere presentare l'esperimento qualche premessa biologica č
necessaria: si tratta, infatti, di organismi dallo stile di vita perlomeno bizzarro,
e che hanno costituito un grosso problema dal punto di vista della loro classificazione
evoluzionistica: ancora recentemente si raggruppava in un unico gruppo "Myxomicetes"
(in origine considerato un sottogruppo dei funghi, poi spostato tra i protozoi:
e questa classificazione č ancora quella proposta nello schema generale
del Brock che vi ho riprodotto) tutti i cosiddetti "slime molds", poi si sono divisi
i "myxomyceti cellulari" (Dictyostelides e Acrasiales) dai "myxomyceti
acellulari (propri)", per giungere in anni recentissimi (in base a seqenze dei 16s rRNA)
ad una completa ridistribuzione in almeno tre taxa distinti, uno per i "vecchi"
myxomyceti propri (o perlomeno per il loro ordine Physarales, l'unico finora
completamente sequenziato), uno per le Acrasiales ed un
altro per i Dictyostelides, l'altro gruppo di myxomyceti cellulari, morfologicamente e
metabolicamente (ma non genicamente) assai simili. Molte specie, inoltre, sono state
spostate da un gruppo all'altro, specie tra acrasiali e dictyostelidi, tra cui anche
il Dictyostelium discoideum specialmente studiato da Bonner. L'ipotesi piů recente sulla
posizione filogenetica delle acrasie č la seguente:
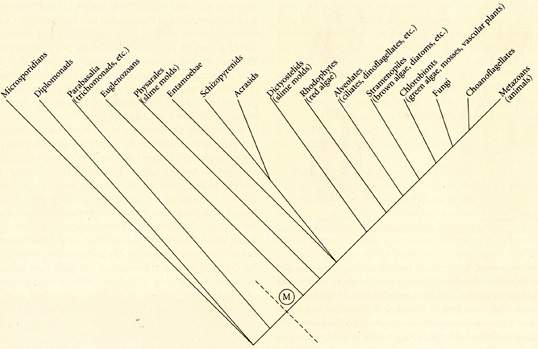
[tav. 8]
Possibile albero cladistico della filogenesi degli eucarioti. Riprodotto da Douglas
J. Futuyuma, Evolutionary Biology, 3rd edition, Sinauer
Associates, Sunderland (Massachusetts), 1998, p. 173.
Si noti, nello schema sopra riportato la collocazione molto
"alta" di alcuni taxa di protozoi (quelli che una volta consideravamo "animali unicellulari")
come i ciliati rispetto ai myxomyceti in genere: stiamo pertanto davvero esaminando una
forma relativamente poco evoluta di eucarioti.
Per comprendere meglio l'esperimento dobbiamo comprendere almeno sommariamente
lo stile di vita di questi particolari myxomiceti, che a volte si comportano come
funghi (formazione di corpi fruttiferi ancorati al suolo e sporazione), a volte come
protozoi (myxoamoebe, cellule libere) ed a volte come "animali" (plasmodii, aggregati
viscidi che si spostano e si nutrono cammin facendo). Lo schema seguente
riproduce il ciclo vitale dei dyctiostelidi (quello delle acrasie, almeno per la
parte che ci riguarda, č molto simile - ma purtroppo non ne sono riuscito a trovare
un disegno altrettanto efficace), puň essere abbastanza chiarificatore:
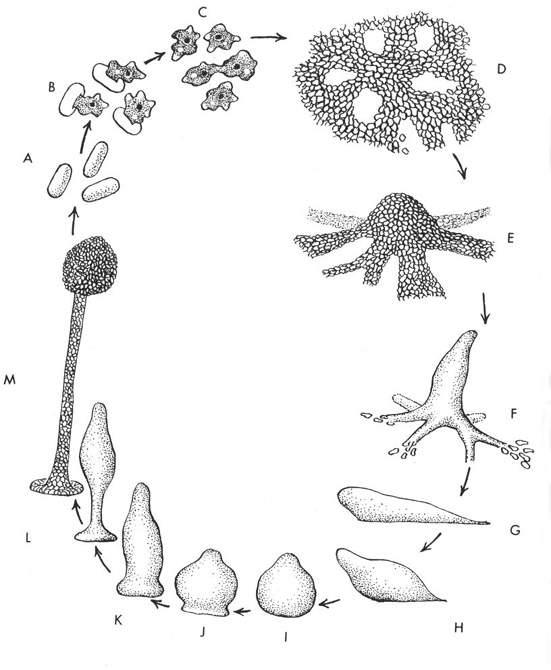
[tav. 9]
Le fasi sono le seguenti: A spore, B germinazione delle spore, C myxoamoebe, D-F
aggregazione cellulare, G Pseudoplasmodio, H-L formazione del corpo fruttifero, M
corpo fruttifero (sorocarpo) maturo, M-A produzione spore e ripetizione del ciclo.
Riprodotto da Steven L. Stephenson & Henry Stempen, Myxomycetes. A Handbook of Slime Molds,
Portland (Oregon), Timber Press, 1994, p. 20.
Come fanno le myxoamoebe ad aggregarsi (fasi D-F nella tavola prec.)
a formare lo pseudoplasmodio? Č appunto in questa fase del ciclo vitale delle acrasie
che avviene la "comunicazione". «In breve, - riporto le parole di Sebeok, Zoosemiotica,
cit., p. 242 - l'aggregazione si verifica in questo modo: un'ameba libera un composto chimico
chiamato da Bonner acrasina [...] che provoca la liberazione di altra acrasina da parte
di amebe molto vicine, dando impulsi a degli impulsi centrifughi, ad onda, della sostanza.
L'acrasina non solo induce la liberazione di altra acrasina, ma fa anche fluttuare
le amebe verso il produttore centrale, formando cosě l'aggregazione». L'importanza non
č tanto nel fatto che le myxoamoebe si "rispondono" a colpi di acrasina ("a chemotactically
active substance", secondo la definizione del Ainsworth & Bisby's Dictionary of Fungi, 8th
edition prepared by the International Mycological Institute, Wallingford (UK), CAB
International, 1995, p. 4), cosa che potrebbe essere una pura reazione chimica condizionata,
quanto nel fatto che esse seguono il gradiente di concentrazione di acrasina nel mezzo
in cui sono flottanti in relazione al primo momento di emissione delle sostanza (in altre
parole che sanno attribuire un preciso valore semantico ai segnali di cui sono
riceventi ed emittenti):
«non č il gradiente "per sé" ad orientare le amebe; č invece la sequenza di tempo
in cui l'ameba emittente produce acrasina. In breve, la struttura dell'impulso orienta
l'ameba, ed č probabilmente specifica» (ibidem).
Č difficile non vedere nel caso che abbiamo descritto una situazione
di reale comunicazione linguistica, anche se il canale (chimico, ed in secondo grado
di potenziale elettrico) di cui essa si serve č radicalmente diverso da quello (fonico)
della comunicazione umana. D'altra parte avevamo giŕ visto come, partendo dal modello
segnico di Jakobson, fosse stata evidenziata la esistenza di tutta una molteplicitŕ
di canali possibili, alternanti o coesistenti (cfr. la tavola
che riproducevamo da Sebeok).
1.5.6 Il "linguaggio" dei procarioti.
Fenomeni analoghi a quelli descritti per le acrasiali (anche se
spesso meno chiari da interpretare, o piů difficili da ottenere) sono riscontrabili
anche altrove, tanto che si potrebbe parlare del canale chimico come del normale
canale della comunicazione cellulare (naturalmente, anche nei neuroni, che sono
delle specialissime cellule, del nostro sistema nervoso lo scambio di informazioni
che avviene nelle sinapsi avviene tramite un meccanismo ionico, ossia elettrochimico).
Una volta che abbiamo accettato la "particolaritŕ" del "linguaggio chimico" delle
cellule, anzi, il suo reperimento in organismi ancora piů primitivi degli eucarioti
ci stupirŕ meno.
I procarioti (batteri ed archeobatteri: cfr la filogenesi
generale del vivente) sono diversi dai procarioti per vari aspetti, tra cui quello per noi
piů importante č che sono quasi del tutto privi di strutture specializzate, che invece
anche gli unicellulari eucarioti avevano: tutti i processi vitali della cellula,
dal metabolismo alla riproduzione, vengono nei procarioti svolti indifferenziatamente
in qualsiasi zona del citoplasma, laddove nei procarioti vengono svolti in strutture
specializzate (reticolo endoplasmatico nel primo caso, e nucleo nel secondo, ad esempio).
Il problema cui dovremo far fronte sarŕ, pertanto, piuttosto spiegare l'assenza di una struttura
specializzata ("memoria" od altro) che possa fungere da supporto al linguaggio, e spiegare
la natura speciale che tale linguaggio dovrŕ avere, perché, di fatto, ormai sappiamo
con certezza che anche i batteri comunicano.
Non sto a riportare altri esperimenti al riguardo, essendomi giŕ
molto diffuso sul "linguaggio" delle acrasie. Basti dire che i biologi sono talmente
sicuri riguardo al linguaggio dei batteri che propongono tra i cinque moderni criteri
con cui identificare il vivente anche proprio la comunicazione:
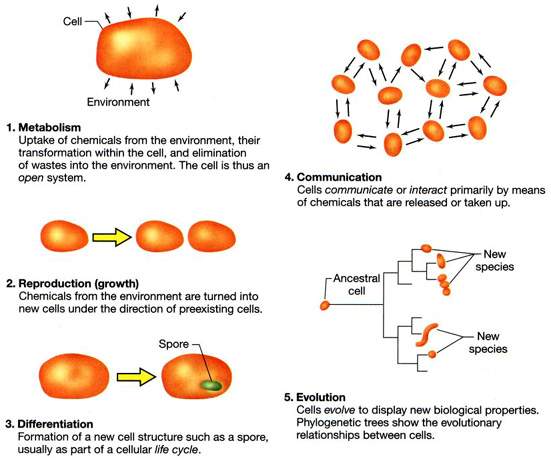
[tav. 10]
Le caratteristiche distintive della vita cellulare. Riprodotto da Brock. Biology
of Microorganisms, 9th edition by M. T. Madigan, J. M. Martinko and J. Parker,
Upper Saddle River (New Jersey), Prentice-Hall, 2000, p. 5.
E tale sicurezza č condivisa dalla tradizione (bio)semiotica:
Sebeok, giŕ nella definizione iniziale della sua Introduction to Semiotics
asseriva che «the phenomenon that distinguishes life forms from inanimate objects
is semiosis. This can be defined simply as the instinctive capacity of all
living organisms to produce and understand signs» (Thomas A. Sebeok, Signs:
An Introduction to Semiotics. Second Edition, Toronto - Buffalo - London, University
of Toronto Press, 2001 [1994 1st ed.], p. 3).
Dando per scontato che i batteri comunichino, e che il canale
sia quello chimico, potremmo perň porre piů precisamente il problema delle strutture
di supporto al linguaggio: anche se le cellule procariote non sono cellule specializzate, devono
pur avere delle strutture fisiche per "esprimere" chimicamente ("parole") il "contenuto" dei loro
"atti comunicativi" (1), il cui "significato" e la cui "codifica" ("langue") deve essere ben
immagazzinata (2) in qualche sorta di "memoria" del batterio.
Una cellula batterica non contiene molto, parete cellulare, citoplasma
ribosomi (RNA e proteine), genoma (DNA e plasmidi), sicché non c'č molto da sciegliere.
La risposta al primo quesito č la piů semplice. L' "apparato articolatorio" del linguaggio
chimico del batterio č, praticamente, la parete cellulare medesima, la cui struttura
č molto complessa: ve ne sono di due tipi fondamentali (gruppo "gram-positivi":
membrana + spesso strato rigido; gruppo "gram-negativi": membrana interna + strato
rigido + membrana esterna), nella tavola seguente rappresento il tipo cui appartiene
il piů studiato dei batteri, l' Escherichia coli:
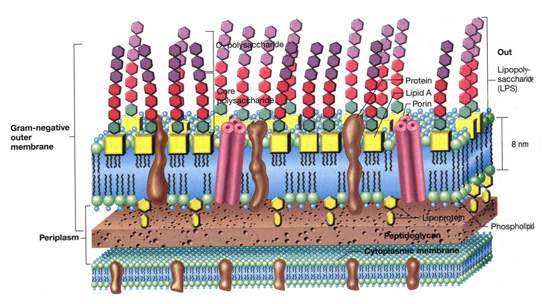
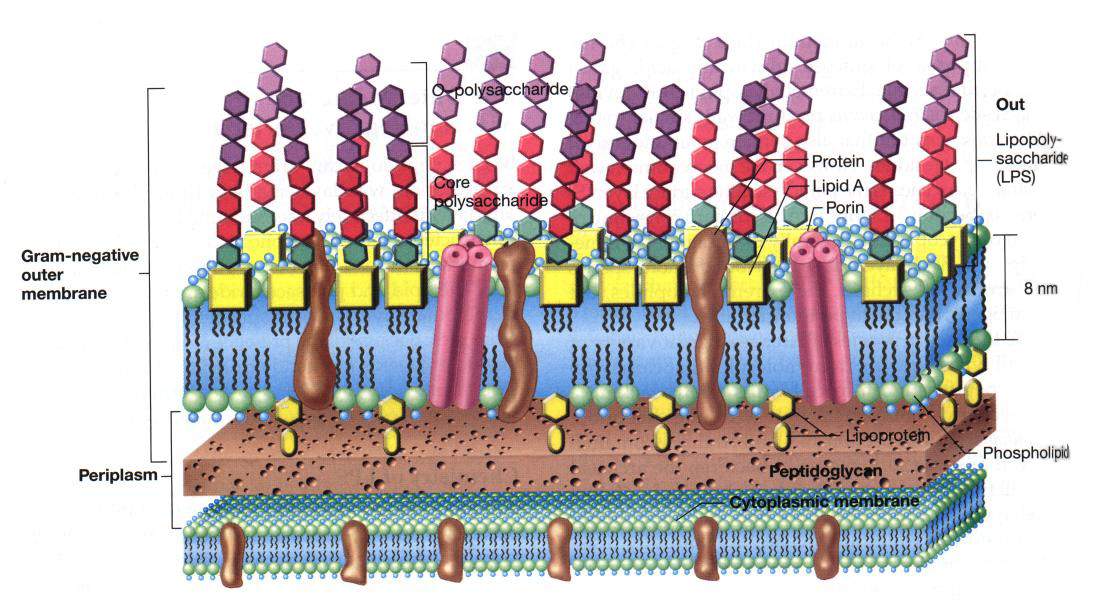
[tav. 11]
La struttura della parete cellulare di un batterio gram-negativo: La parete cellulare dei gram-negativi.
La membrana interna o citoplasmatica č costituita da un doppio strato di fosfolipidi
(alla stessa maniera della membrana dei gram-positivi e degli eucarioti), lo strato
rigido č costituito da un fitto reticolo di peptidoglicani (sostanza assente dalla membrana
cellulare eucariota), mentre la membrana esterna, fondamentalmente un bistrato di
fosfolipidi anch'essa, ha una complessa impalcatura di molecole
che si protendono all'esterno (a modo di "sensori") e di proteine (tra cui le porine) che
permettono il passaggio di molecole o cariche ioniche tra l'interno e l'esterno della
cellula. Riprodotto da Brock. Biology of Microorganisms, 9th edition
by M. T. Madigan, J. M. Martinko and J. Parker, Upper Saddle River (New Jersey),
Prentice-Hall, 2000.
Il funzionamento della membrana č molto complesso, ma, se accettiamo
di non addentrarci nel dettaglio chimico del suo funzionamento, concettualmente č
abbastanza comprensibile. Una sfida maggiore č posta dal secondo problema: dove risieda
la "memoria" del batterio, tanto quella operativa a breve termine, quanto quella
a lungo termine dove sarŕ depositata la sua "langue", il suo "lessico". I batteri,
in effetti, presentano entrambi i tipi di fenomeni.
Il corrispondente della memoria
operativa, con cui il batterio puň rispondere rapidamente alle sollecitazione esterne,
č occupato da un gruppo di complesse reazioni chimiche (troppo complicate perché
le possa qui spiegare; cfr. comunque Joseph W. Lengeler - Gerhart Drews - Hans G.
Schlegel, Biology of the Prokaryotes, Stuttgart - New York, Georg Thieme Verlag,
1999, pp. 469-490) chiamato posttranslational control (legato ai
ribosomi dove avviene la "traduzione" del RNA in proteine), grazie al quale le
proteine fabbricate dal batterio possono venire adattate conformemente
agli stimoli esterni per poi venire "trasportate" a rispondervi (diventando, per modo
di dire, le sue "paroles").
La risposta a quale sia invece il corrispondente della cruciale memoria a lungo termine
č, invece, al contempo semplice e paradossale: nel genoma (DNA, RNA e proteine non a caso
sono di solito definite come "macromolecole informazionali" proprio perché contengono
informazioni biologiche nelle loro sequenza). L'unica memoria possibile per depositare
la semantica (che puň minimalmente essere anche solo il valore per la sopravvivenza del
batterio) di cose (sostanze chimiche, altri batteri, virus) con cui il batterio entra
in contatto č quindi la modificazione del corredo genico. Paradossalmente, se la conoscenza
del batterio č SOLO codificata genicamente, il batterio sarebbe l'unico organismo
il cui linguaggio sia integralisticamente chomskyano (nonostante per Chomsky non si
possa parlare di linguaggio fuori dall'uomo), ed anzi anche completamente platonico
(la conoscenza attinge solo al ricordo di vite precedenti ...).
La cosa non č senza conseguenze apparentemente gravissime: la natura rigorosamente
genetica della sua "memoria", infatti, comporta che il batterio non puň "imparare"
come individuo ma solo come specie. Innoltre il genoma non si modifica certo
direttamente: tranne che nel caso delle alterazioni apportate da un virus o dalla
reazione antigenica ad esso, ogni mutamento deve avvenire tramite il lento processo
della selezione naturale. E questo, rapportato al passo degli eucarioti superiori,
equivalerebbe a dire che ci vorrebbero milioni di anni perché una nuova conoscenza
venga memorizzata secondo questo sistema. Tutto ciň sembrerebbe oltremodo poco funzionale:
potrŕ mai funzionare? A vedere i fatti parrebbe proprio di sě: anzi, il meccanismo č
talmente ben adattato che i piů antichi fossili risalgono a 3.600 milioni di anni fa,
e non sembtrano molto diversi dai batteri odierni:
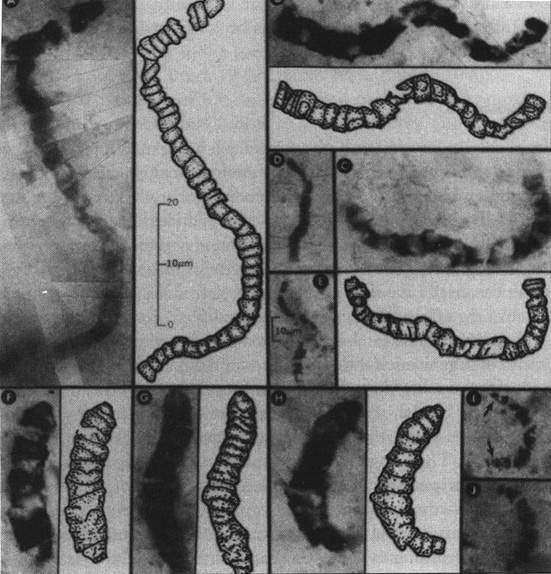
[tav. 12]
I primi batteri fossili, risalenti a 3.600 milioni di anni fa. Da J. William Schops,
Microfossils of the Early Archean Apex chert: New Evidence of the Antiquity of Life,
in "Science" CCLX (1993) 620-646; riprodotto anche in Ernst Mayr, What Evolution Is,
New York, Basic Books, 2001. Altre stime variano di pco la cronologia, riportandoci a 3,86
bilioni.
E considerate che 3,6 ~ 3,8 bilioni di anni fa sono proprio tanti: la vita stessa
appare "solo" cento milioni di anni prima, circa 4,6 bilioni di anni fa:

[tav. 13]
Riprodotto da Ernst Mayr, What Evolution Is, New York,
Basic Books, 2001.
La spiegazione dell'apparente aporia risiede in una caratteristica
del sistema di riproduzione dei procarioti che, associata alla rapiditŕ media dei cicli
loro metabolici, fa sě che le mutazioni geniche si fissino in tempi abbastanza rapidi
(vedi ad esempio la resistenza agli antibiotici sviluppata da molti batteri nel giro
di poche generazioni umane). Negli eucarioti, infatti, la percentuale di errore ammmessa
nella replicazione del DNA č molto bassa (c. 10-11), grazie anche alla notevole efficienza
raggiunta da un meccanismo noto come "proof reading" ('correzione di bozze') che riparara la
maggior parte degli errori. Nei procarioti tale sistema č meno efficiente e la percentuale
di errorre ammessa sale ad intorno al 10-7 (cfr. Joseph W. Lengeler - Gerhart Drews -
Hans G. Schlegel, Biology of the Prokaryotes, Stuttgart - New York, Georg Thieme Verlag,
1999, pp. 376-384): apparentemente questa strategia č controproducente per la
stabilitŕ della specie, ma costituisce anche un grande vantaggio evolutivo. E rende
meno inefficiente anche la fissazione genetica dell'informazione nelle basi del genoma.
1.5.7 Biologia e biosemiotica: origine del linguaggio e della vita.
Inseguendo il linguaggio lungo le linee evoluzionistiche del vivente
ci siamo ormai avvicinati molto al problema stesso dell'origine della vita. Forse che
linguaggio e vita siano due "stranezze" legate insieme fin dall'inizio?. Alcuni indizi
farebbero pensare di sě.
Una delle teorie recenti piů accreditate sull'origine della vita č la cosiddetta teoria
del "mondo del RNA", per cui cfr. la tavola qui sotto. Ipersemplificando, nel
brodo primordiale delle prime molecole pre-organiche il passaggio dallo stato inorganico
(le sostanze, insiemi di molecole dello stesso tipo, si trasformano non direzionalmente, ma
solo in base a reazioni chimiche meccaniche e non finalizzate), attraverso vari passaggi
intermedi (la formazione dei "cristalli", ad esempio, rappresenta giŕ un grado di
organizzazione delle molecole di una sostanza piů finalizzato), a quello organico (una molecola
reagisce chimicamente in modo finalizzato, ossia si "riproduce") sarebbe avvenuto
inizialmente in molecole di RNA. La replicazione di una molecola di acido nucleico viene
normalmente promossa da una proteina ( che agisce come "enzima") esterno: sicché il problema (viene prima l'acido
nucleico o la proteina?) sembrava configurarsi come una riproposizione del vecchio paradosso
"č nato prima l'uovo o la gallina?". Si sono perň accumulate molte prove che l' RNA
puň anche agire come un enzima: piů precisamente, alcune sequenze di basi del RNA,
le "ribozyme", agiscono come enzima, tagliando, dividendo e prolungando altri
nucleotidi. Inoltre, č stato dimostrato che brevi sequenze modello di RNA possono autocatalizzare
la formazione di sequenze complementari a partire da nucleotidi liberi ("autocatalisi"). Naturalmente
questo presuppone la compresenza di una quantitŕ di sequenze di RNA e nucleotidi liberi
in uno stesso sistema chiuso, che agirebbero nel modo rappresentato nella tavola seguente:
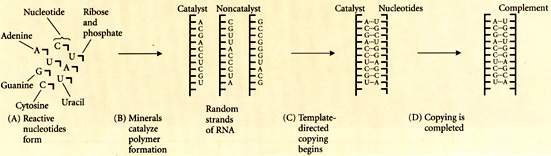
[tav. 14]
L'origine della vita: l'ipotesi del "mondo del RNA". Riprodotto da Douglas J. Futuyuma,
Evolutionary Biology, 3rd edition, Sinauer Associates, Sunderland (Massachusetts),
1998, p. 173. La mia esposizione semplificata della teoria č basata, oltre che su Futuyama,
cit. anche su John Maynard Smiyh - Eörs Szatmháry, The Major Transitions in Evolution,
Oxford, Oxford University Press, 1995.
Per il nostro discorso č importante constatare che, perché una sequenza
di RNA (ribozyma) possa fare da "codice" per la replicazione (informazione codificata),
č indispensabile la presenza concomitante di tutta una pluralitŕ di molecole: che č precisamente il rapporto
necessario tra "significazione" e "popolazione" configurato nella "convenzionalitŕ" saussuriana.
Sembra di intravedere in modo aurorale qualcosa di molto simile a quella "convenzione" tra
appartenenti ad una comunitŕ linguistica che avevamo individuato come presupposto "logico" (anche se non
necessariamente "storico") della esistenza di un linguaggio. Sembra quasi, in altre
parole, che la nascita della vita abbia le stesse condizioni della nascita del linguaggio.
In questo senso pensare che la significazione linguistica (e proprio nel senso di Saussure!)
sia da legare indissolubilmente alla nascita della vita non č forse una idea troppo
ardita: ed č questa, propriamente, la tesi della biosemiotica.
1.5.8 Per una tipologia dei linguaggi.
Ho lasciato a questo ultimo paragrafo il compito di contenere
la tabella, cui abbiamo fatto costante riferimento in tutto il capitolo, in cui abbiamo
provato ad abbozzare una tipologia riassuntiva dei vari tipi di linguaggio. Le prerogative
sono segnalate in modo solo approssimativo, e si intende comunque che vadano integrate
e precisate con quanto abbiamo detto piů distesamente prima:
| Linguaggio |
| linguaggi naturali | linguaggi artificiali |
| linguaggio umano | linguaggi animali | linguaggi logici | linguaggi informatici | lingue inventate (esperanto) | metaling. funzionali (ASL) | codici di segnali |
| (scimpanzé) bonobo | cnidaria ... eucaria | procaria |
| segno linguistico | + | + | ---- | (±) | (+) | (–) | + | (+) | – |
| discretezza | articolazione | + | (±) | ---- | (–) | (+) | (±) | + | ± | (+) |
| composizionalitŕ | + | (+) | (+) | (±) | + | (+) | + | (±) | (–) |
| ricorsivitŕ | + | (±) | ---- | – | + | + | + | (+) | (–) |
| dipendenza dalla struttura | + | (±) | ---- | – | (±) | (–) | (+) | (±) | (–) |
| storicitŕ | lingue storiche | + | (±) | ---- | ? | + | + | + | + | + |
| diacronia | + | ? | ---- | ? | – | (–) | ± | ± | ± |
| socialitŕ - convenzionalitŕ | + | + | ---- | (±) | + | + | + | + | + |
| strutture specializzate | + | + | ± | (–) | x | + | x | x | x |
| acquisizione naturale | + | (+) | ---- | ? | – | – | – | – | – |
| uso spontaneo | + | (+) | ---- | ? | – | – | (–) | (–) | – |
[tav. 15]
Una tipologia delle principali caratteristiche del linguaggio, umano e non umano.
{kind=link}
{kind=link}
{kind=link}
{kind=link}